
ミトコンドリア : 細胞シグナル伝達とエネルギーバランスの中心
ミトコンドリアは、内膜 / 外膜、膜間腔、クリステ、マトリックスから構成され、核DNAとは別のDNAを含みます。
ミトコンドリアは、内膜と外膜、膜間腔、クリステ、およびマトリックスから構成されており、核DNAから分離された独自のDNAを内包しています。タンパク質の細胞内位置に関する知識は、多様な細胞プロセスにおいてそのタンパク質が果たす潜在的役割を明らかにする可能性があります。ここにリスト化されているオルガネラ特異的マーカーの一つとの共局在化により、目的とするミトコンドリアタンパク質の細胞内位置を確認することができます。
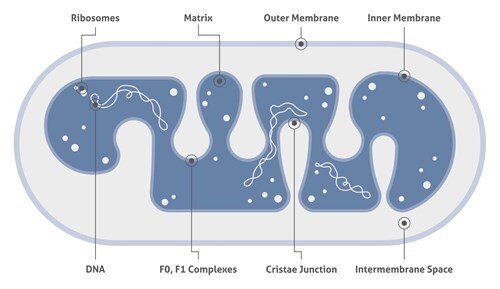
クエン酸回路
クエン酸回路(トリカルボン酸(TCA)回路またはクレブス回路としても知られている)は、すべての好気性生物が使用する一連の化学反応であり、炭水化物、脂肪、およびタンパク質から誘導された酢酸エステルの酸化を介して、二酸化炭素と、アデノシン三リン酸(ATP)の形の化学エネルギーを生成します。加えて、このサイクルにより、特定のアミノ酸の前駆体、ならびに多くの他の生化学反応で使用される還元剤であるNADHが生成されます。
真核細胞では、クエン酸回路の反応は、ミトコンドリアのマトリックス中で行われます。回路の反応は8つの酵素によって行われ、アセチルCoAの形の酢酸エステルを水と2分子の二酸化炭素まで完全に酸化します。
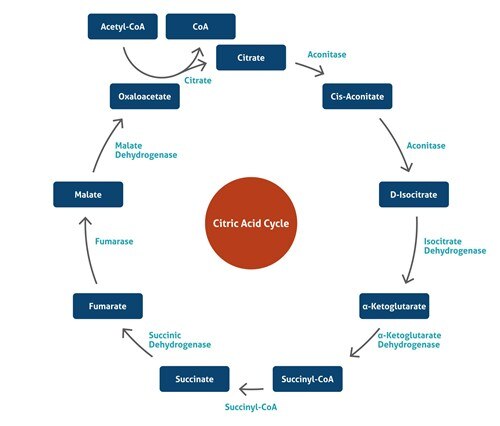
ミトコンドリア呼吸複合体
細胞呼吸は、食物からエネルギーを放出し、生命過程のためのエネルギーを供給する過程です。ミトコンドリア呼吸鎖は細胞呼吸の最終的で最も重要な段階で、ミトコンドリアの内膜上に位置し、4つの大きな膜貫通タンパク質複合体(呼吸鎖複合体I、II、III、IV)(CI、CII、CIII、CIV)のほか、CI/IIとIIIの間のユビキノンとCIIIとIVの間のチトクロームcからなります。ミトコンドリアの呼吸鎖の機能は、NADHとコハク酸から電子を酸素に伝達し、次に内膜を横切ってプロトン濃度勾配をつくることによる生物学的酸化です。
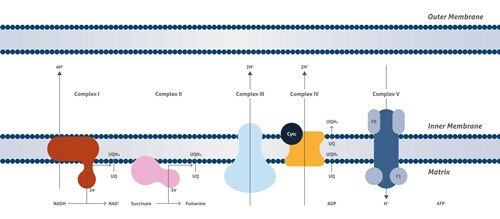
ミトコンドリアの分裂
ミトコンドリアは、細胞機能に不可欠であり、真核生物のエネルギー生成、代謝物、リン脂質、およびヘムの合成、ならびにカルシウム恒常性の維持を担っています。ミトコンドリアは、極めてダイナミックなオルガネラであり、融合および分裂イベントを高頻度で行っています。融合と分裂という反対のプロセスによってミトコンドリアの形態は維持されており、mtDNAと代謝混合、生体エネルギー機能、およびオルガネラ数の維持を確実にするのはこの平衡です。
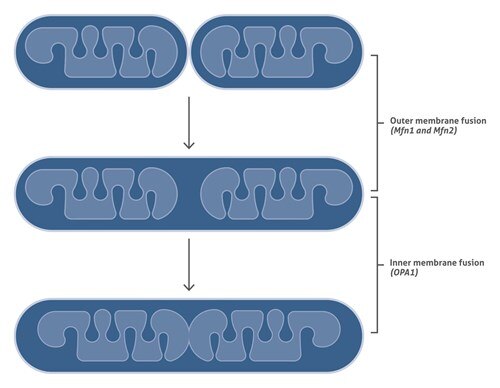
ミトコンドリアの融合
ミトコンドリアの融合は3段階で起こります: 1) ミトコンドリアの外膜を介して2つのミトコンドリアがドッキング、2)外膜の融合、および3)内膜の融合です。最初の2つの段階は、ミトフュージョンタンパク質(哺乳類におけるMfn1およびMfn2)により仲介されます。その次の段階(ミトコンドリア内膜の融合)は、ダイナミン関連GTPase OPA1により仲介されます。
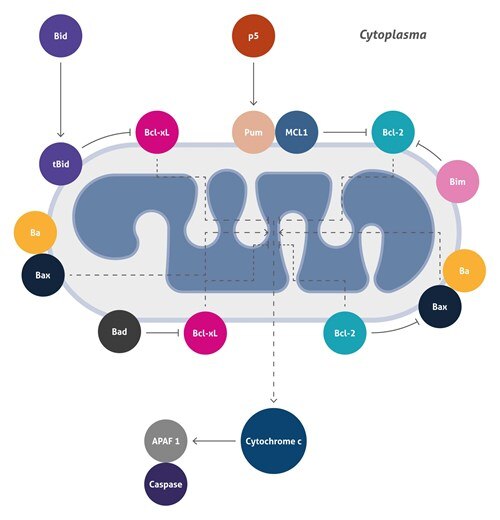
ミトコンドリア仲介性アポトーシス
アポトーシスは、プログラムされた細胞死(PCD)のプロセスであり、動物の発生および組織の恒常性において中心的役割を果たします。今日までに知られている2つの主要なアポトーシス経路があり、ミトコンドリア(「内因性」経路)または細胞表面受容体(「外因性」経路)のいずれかにより開始されます。ミトコンドリア仲介によるアポトーシスは、腫瘍抑制タンパク質(例えばp53)および癌遺伝子(例えばc-Myc)の活性化、DNA損傷、化学療法薬、血清飢餓、および紫外線照射など、幅広い細胞刺激に応答して起こります。この経路は、細胞内部で始まり、ミトコンドリア透過性が亢進して、アポトーシス促進分子であるシトクロムcの細胞質への放出が起こります。
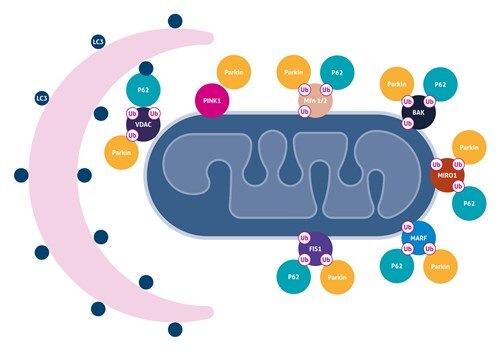
ミトコンドリアオートファジー
ミトコンドリアオートファジーは、オートファゴソームによる損傷したミトコンドリアの選択的除去と、それに続くリソソームによる異化のプロセスです。1つの一般的なメカニズムは、ミトコンドリアの脱分極がPINK1の安定化とミトコンドリアへのパーキンの動員をもたらすものです。
パーキンは、ミトフシン1および2(MFN1および2)、ヘキソキナーゼ、TOM複合体成分、FIS1、BAK、MIRO、ならびにVDACをユビキチン化することができます。これらは、プロテアソームを介して分解されるか、p62の結合パートナーとして機能します。p62は、LC3との直接相互作用を介してアダプター分子として機能し、オートファゴソーム膜をミトコンドリアに動員することができます。パーキンはAmbra1とも相互作用することができ、Ambra1はミトコンドリア周辺のPI3K複合体を活性化して、選択的マイトファジーを促進します。
ミトコンドリア翻訳
ミトコンドリアには、核から分離された独自のDNAがあります。進化の過程で、ミトコンドリアのタンパク質コード遺伝子は、ほとんどが核ゲノムに移されました。しかし、少数の遺伝子は現代のオルガネラのゲノムに保持されています。
ミトコンドリアマトリックスに収容されているミトコンドリアゲノム(mtDNA)には、13のタンパク質の青写真と、ミトコンドリア内タンパク質合成に必要かつ十分であると考えられているRNA分子がすべて含まれていることが現時点で分かっています。ミトコンドリア内タンパク質合成に必要な他の成分は、すべて細胞質で合成された後にサイトゾルから取り込まれます。ミトコンドリアタンパク質合成のプロセスの中心は、ミトコンドリアリボソームです。
ミトコンドリアタンパク質の取り込み
ミトコンドリアは、独自のゲノムと翻訳機構を持っていますが、mtDNAによってコードされ、オルガネラ内で合成されるミトコンドリアタンパク質は、呼吸鎖複合体の少数のコア構成要素など少数だけです。したがって、ミトコンドリアタンパク質の大部分は核にコードされており、オルガネラ内に取り込まなければなりません。
遊離リボソーム上で合成されたあとで、ミトコンドリア前駆体タンパク質は、細胞質シャペロンによってガイドされるプロセスにおいてオルガネラの表面に到達します。その後、ミトコンドリア前駆体タンパク質は、特殊なタンパク質取り込み機構によって取り込まれ、外膜、内膜、膜間腔、またはマトリックス内の指定されたミトコンドリア内目的部位に局在化されます。
J R Friedman, J Nunnari. Mitochondrial form and function. Nature. 2014 Jan 16;505(7483):335-43.