オートファジー
はじめに
オートファジー(Autophagy)とは、真核生物における保存された細胞内タンパク質の分解経路です。損傷を受けたオルガネラ(細胞小器官)やミスフォールディングタンパク質(誤って折りたたまれたタンパク質)を除去することで、細胞の恒常性維持に寄与します。オートファジーは、細胞の恒常性や、細胞の増殖、発生、分化等の正常な生理的プロセスにおいて重要な役割を果たすと同時に、神経変性疾患、心筋症、自己免疫疾患、がん等の様々な疾患の病態形成にも関連しています。
プロテインテックは、オートファジー研究をサポートする様々な製品を取り揃えています。
目次 |
オートファジー経路
オートファジーは、(1)オートファゴソームの形成、(2)オートファゴソームとリソソームの融合(オートリソソームの形成)、(3)取り込み物質の分解、の3段階で構成される極めて動的なプロセスです。オートファジーは、栄養飢餓、増殖因子によるシグナル伝達、細胞ストレス等の種々のトリガー要因が関係する様々なシグナル伝達経路によって誘導されます。
ATGタンパク質
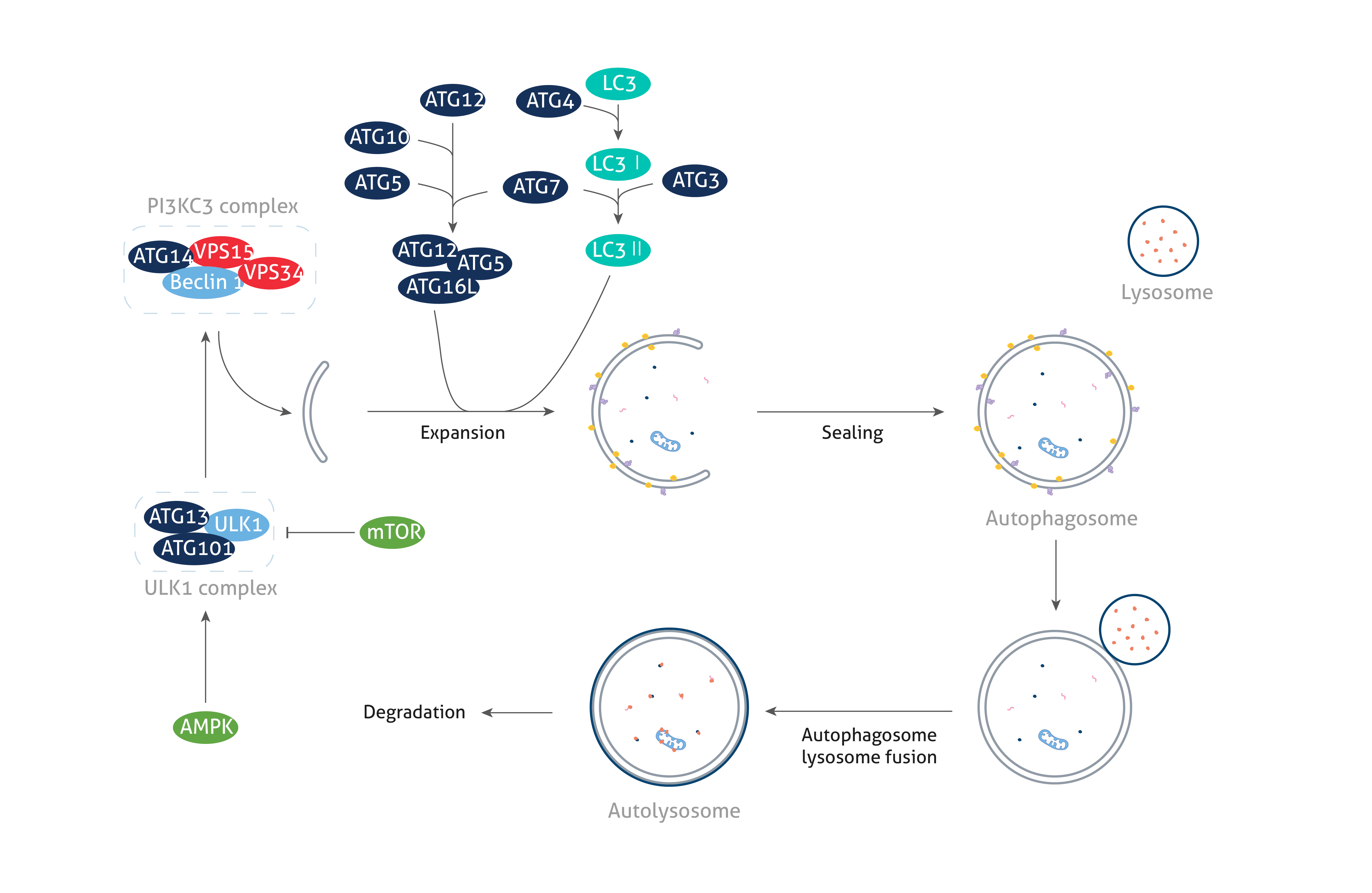
ATG(Autophagy-related、オートファジー関連)タンパク質は、オートファジー経路の非常に重要な特徴である「オートファゴソーム(autophagosome:細胞内成分の一部を隔離して形成される二重膜小胞)」の形成に必須のタンパク質群です。オートファゴソームの形成プロセスは、開始、隔離膜の形成、伸長、閉鎖、最終的なリソソーム融合の過程を経て進行します。各ステップは、様々なATGタンパク質によって制御を受けます。ATGタンパク質群は、オートファゴソーム形成における役割に応じて、次の機能グループに分類することができます。それらは(1)ULK1キナーゼ、(2)クラスⅢPI3K複合体Ⅰ、(3)ATG2-ATG18/WIPI複合体、(4)ATG9輸送(ATG9 trafficking)、(5)ATG5-ATG12結合反応系、(6)ATG8/LC3結合反応系のいずれかに分類されます。
関連製品
| ULK1 | VPS15 | ATG2A | LC3A |
| ATG13 | Beclin 1 | ATG12 | ATG3 |
| RB1CC1 | ATG14 | ATG7 | ATG4A |
| ATG101 | ATG9A | ATG5 | |
| VPS34 | WIPI1 | ATG16L1 |
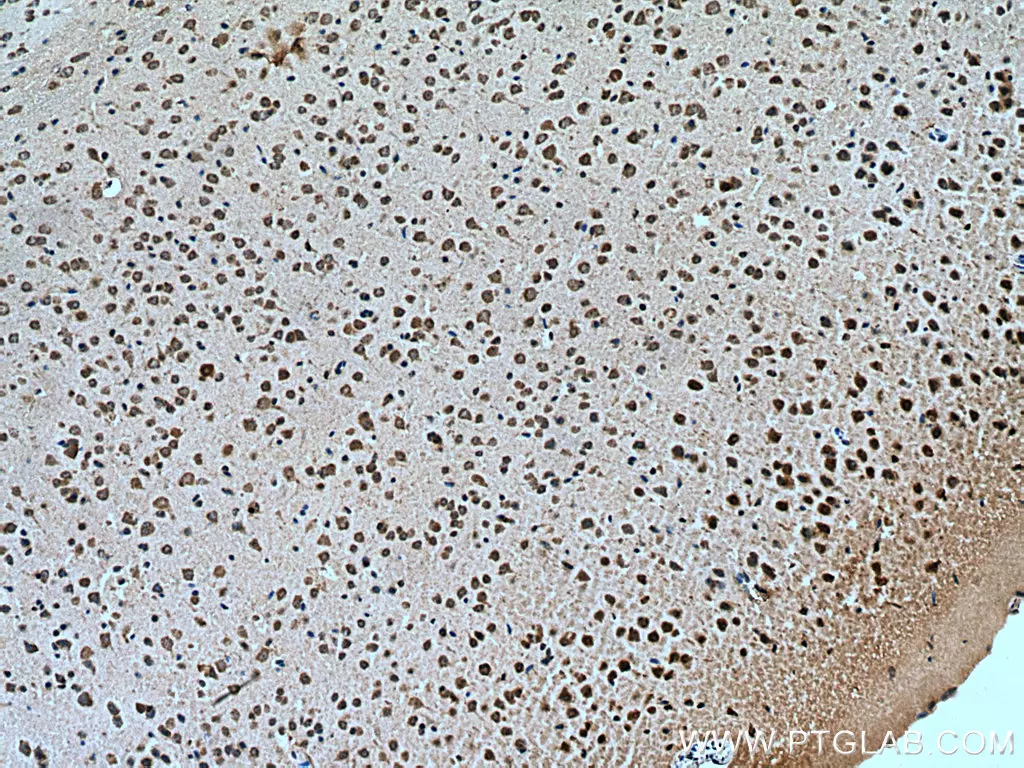
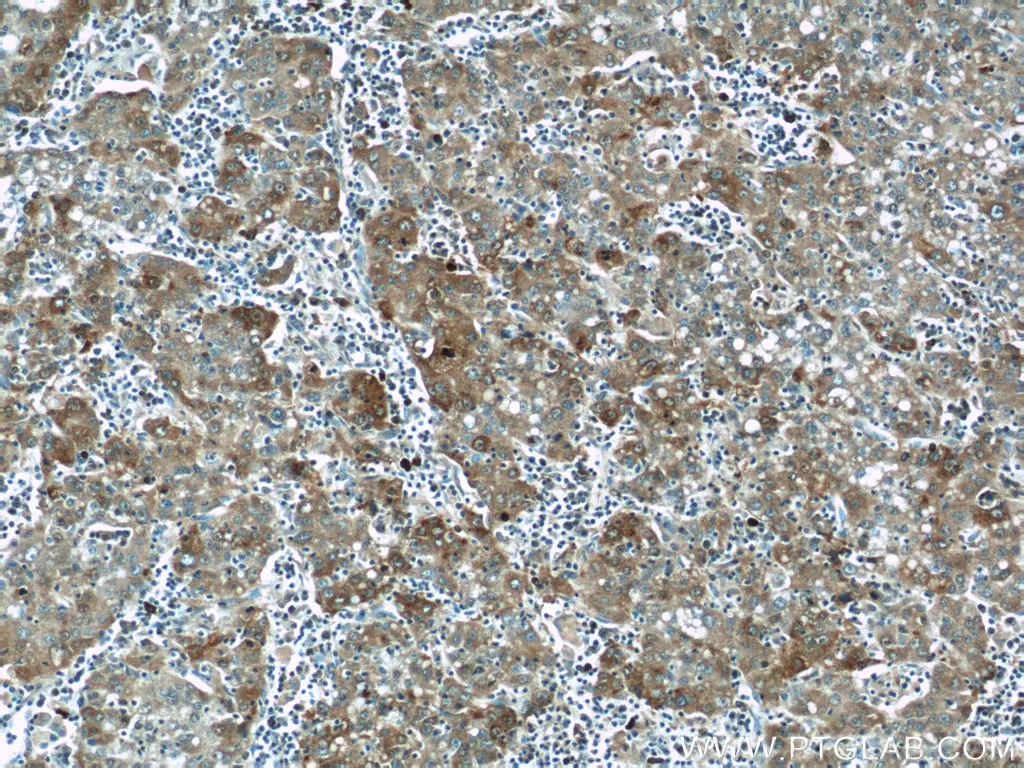
ATG14/Barkor(N-terminal)抗体(カタログ番号:19491-1-AP、希釈倍率1:200)を使用した、パラフィン包埋ヒト肝臓組織の免疫組織化学(IHC:Immunohistochemistry)染色(10倍レンズを使用)。
オートファジーフラックス
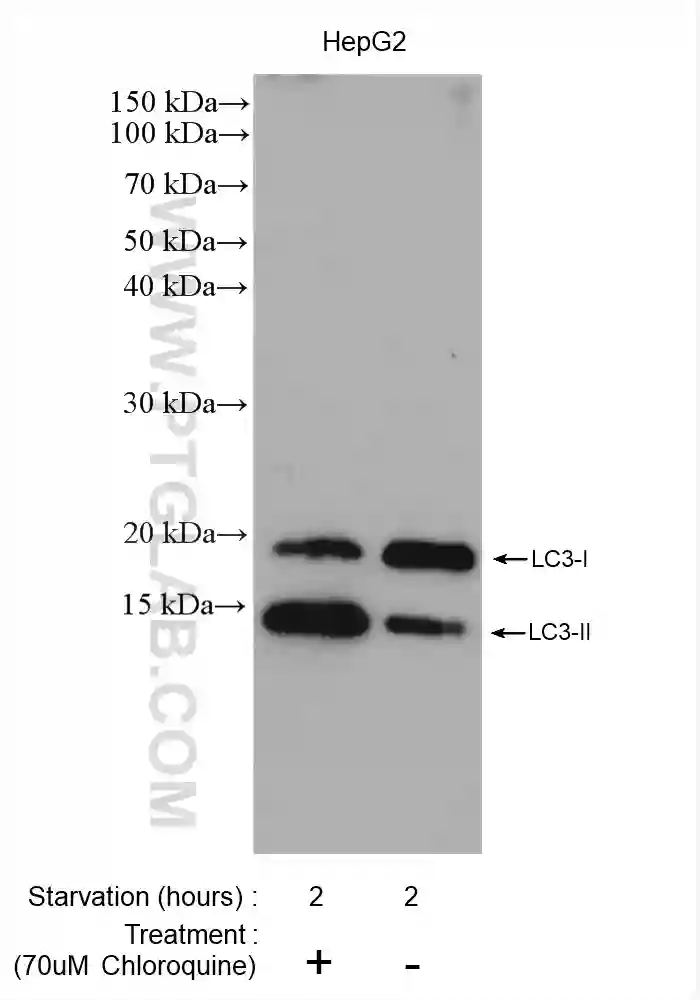
オートファジー活性を計測するための理想的な手段は、オートファジー経路の分解速度を表す「オートファジーフラックス(autophagy flux)」のモニタリングです。オートファジーフラックスの測定に最も幅広く用いられている手法は、ウェスタンブロット(WB:Western blot)によってオートファゴソーム小胞の膜タンパク質であるLC3のプロセシングを検出する手法です。LC3前駆体は、システインプロテアーゼであるATG4によって切断されて、LC3-Ⅰになります。続いてLC3-Ⅰは、ホスファチジルエタノールアミン(PE)と共有結合してLC3-Ⅱになります。オートファゴソームのオートリソソームへの成熟に伴い、リソソームプロテアーゼの作用によって小胞内膜に局在するLC3-Ⅱの分解が生じます。したがって、オートファゴソーム形成の誘導はLC3-Ⅱレベルの増加をもたらすのに対して、オートファゴソームとリソソームの融合(オートリソソームの形成)はLC3-Ⅱレベルの減少をもたらします。
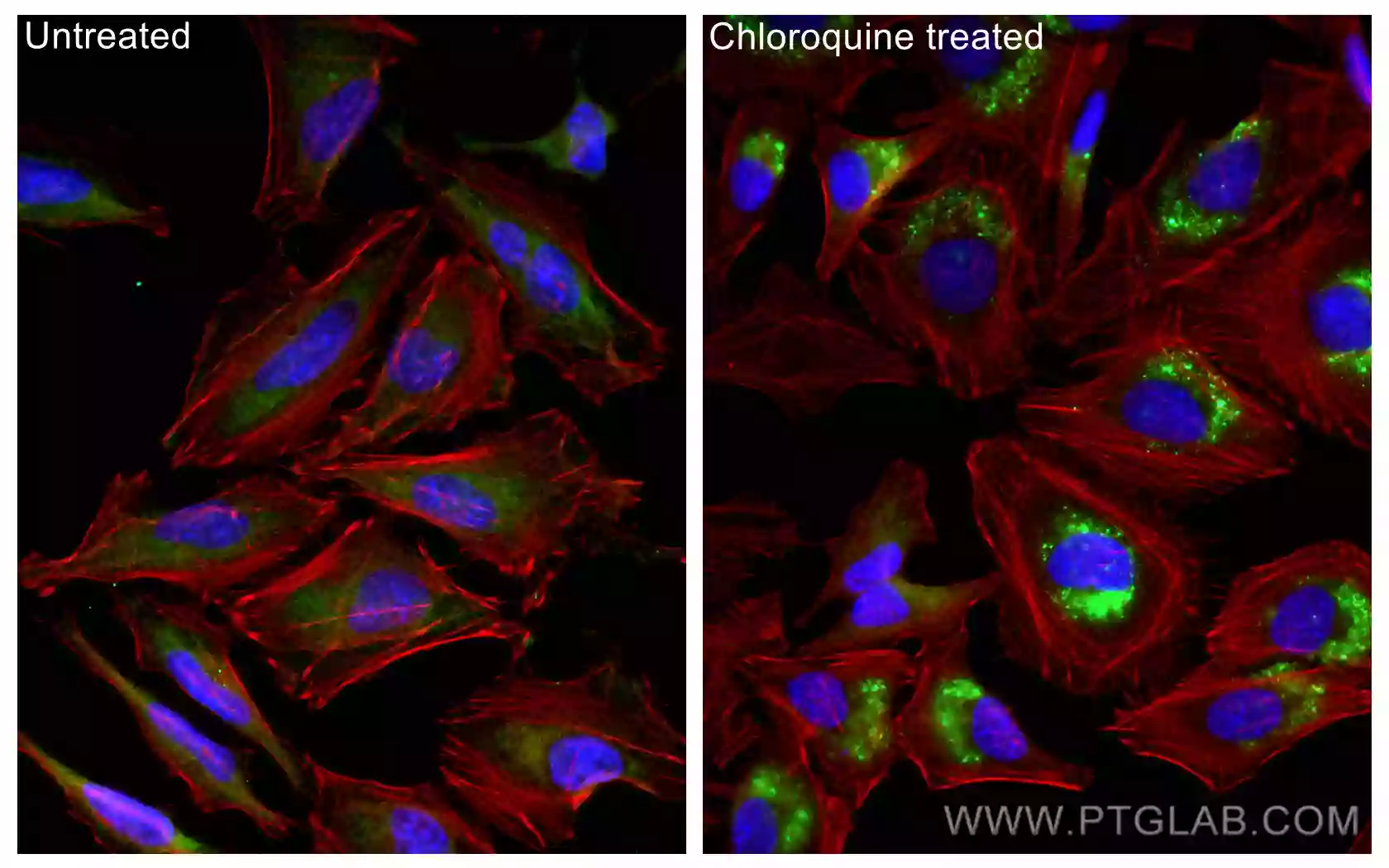
しかし、オートファゴソームとリソソームの融合(オートリソソームの形成)が阻害された場合、LC3-Ⅱ分解の減少に伴ってLC3-Ⅱ蓄積の増加が誘導され、結果としてLC3-Ⅱの増加に寄与することになるため、単にLC3-Ⅱレベルの増加を測定することは、オートファジーフラックスの誘導を評価する最適な手法とはいえない場合があります。そのため、正確に結果を解釈するには、オートファゴソームとリソソームの融合を阻害するリソソーム阻害剤(Chloroquine(クロロキン)等)の存在下および非存在下の両サンプルにおけるLC3-Ⅱ量を測定することが推奨されます。リソソーム阻害剤を添加すると、オートファゴソーム内のLC3-Ⅱのターンオーバーが妨げられます。つまり、リソソーム阻害剤を添加したサンプルにおけるLC3-Ⅱレベルの増加は、実際にオートファジーフラックスが増加したことを示しています。正確にオートファジーフラックスを評価するために、LC3-Ⅱのターンオーバーの測定に加え、オートファジー基質であるp62/SQSTM1等をウェスタンブロット(WB)解析で同時に検出することがしばしば推奨されます。
また、免疫蛍光染色(IF:Immunofluorescence)を実施して、リソソーム阻害剤の存在下および非存在下の両サンプルに存在するLC3数を定量化することで、オートファジーフラックスを解析することも可能です。その他に、オートファゴソームマーカーであるLC3と、リソソームマーカーであるLAMP等を同時に染色して、オートファゴソームとリソソームの融合をモニタリングすることも可能です。
関連製品
| LC3 | p62 | LAMP1 |

オートファジーとミトコンドリア間のクロストーク
オートファジーは、損傷したミトコンドリアの選択的分解に適宜関与することによって、ミトコンドリアの健全性の調節において極めて重要な役割を果たします。このミトコンドリアに対するオートファジー分解プロセスは「マイトファジー(mitophagy)」として知られます。マイトファジーは、ミトコンドリアの膜電位低下が引き金となってPINK1(PTEN induced putative kinase 1)が選択的にミトコンドリアに蓄積し、ユビキチンリガーゼが誘導されることで促進されます。PINK1は、脱分極したミトコンドリアでは代謝回転によって分解されなくなるため安定化し、ミトコンドリア外膜へ蓄積してミトコンドリアにParkinを動員します。Parkinが特定のミトコンドリアタンパク質をユビキチン化すると、ユビキチン化タンパク質にp62が動員されます。その結果、損傷ミトコンドリアはオートファゴソームへ取り込まれて分解されます。マイトファジーは、損傷ミトコンドリアの除去に加えて、NIX(別名:BNIP3L)等のタンパク質によって媒介される過剰なミトコンドリアの除去においても重要な役割を果たします。
オートファジーがミトコンドリアの健全性を調節する一方、ミトコンドリア自体もオートファジー経路に様々な形で影響を与えます。例えば、ミトコンドリアが産生する活性酸素種(ROS:Reactive oxygen species)は、オートファジーの重要な調節因子です。また、ミトコンドリアのATP産生能が低下した場合、ミトコンドリアはmTORやAMPKが媒介するオートファジーを誘導することで、オートファジーフラックスを直接的に調節します。さらに、ミトコンドリアは、飢餓状態によって誘導されるオートファジーにおけるオートファゴソーム膜の潜在的供給源となっている可能性が示唆されています。ミトコンドリアのダイナミクスとして知られる、ミトコンドリアの融合と分裂のバランスの変化も、オートファジーに対する細胞の応答に影響を及ぼします。例えば、ミトコンドリアは飢餓状態に置かれると、オートファジーによる分解から自己を保護するためのメカニズムとして、過剰に融合して伸長することが明らかになっています。
関連製品
| Fis1 | Mfn2 | Parkin |
| Mfn1 | BNIP3L | PINK1 |