
特集:DNA損傷修復マーカー
DNA損傷が修復されないと、突然変異や細胞死の誘発につながります。
DNA変異は、結腸がん、皮膚がん、白血病、リンパ腫、乳がん、卵巣がん等の多くのがんの腫瘍形成の要因としてよく知られています。
筋細胞やニューロン(神経細胞)におけるDNA損傷の蓄積は、変性疾患を引き起こします(1)。ヒトの脳は、体内に取り込んだ酸素の約20%を消費し、その適切な機能を維持するために十分な量のエネルギーを産生します。このエネルギー産生の過程で、ROS(活性酸素種)は自然に生じる副産物であり、脳内で容易に蓄積され、DNA損傷を誘発します。
日々、細胞は様々なDNA損傷因子に曝されています(2)(DNA損傷修復マーカーに関するブログ記事もご覧ください)。ゲノム安定性のセーフガードとして、細胞は、酸化、加水分解、DNA塩基の二量体化(チミン二量体化)、一本鎖/二本鎖切断による損傷に対する修復経路を発達させてきました(図1)。
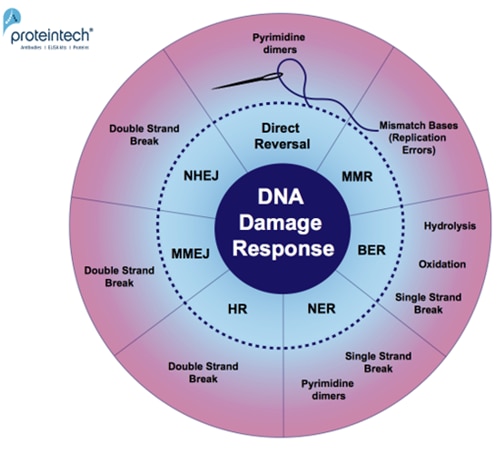
図1. DNA損傷特異的修復経路の概要。応答メカニズム:直接修復(Direct reversal)、MMR(MisMatch Repair:ミスマッチ修復)、BER(Base Excision Repair:塩基除去修復)、NER(Nucleotide Excision Repair:ヌクレオチド除去修復)、HR(Homologous Recombination:相同組換え)、MMEJ(Microhomology-Mediated End Joining:マイクロホモロジー媒介末端結合)、およびNHEJ(Non-Homologous End Joining:非相同末端結合)。
気候変動や環境汚染の増加により、DNA損傷因子に曝される機会は増加しています。そのため、DNA損傷の修復メカニズムを解明することの重要性は益々高まっています。Tomas Lindahl氏、Paul Modrich氏、Aziz Sancar氏らは、DNA損傷修復経路を分子レベルで明らかにした先駆者でした(2015年、ノーベル化学賞)。
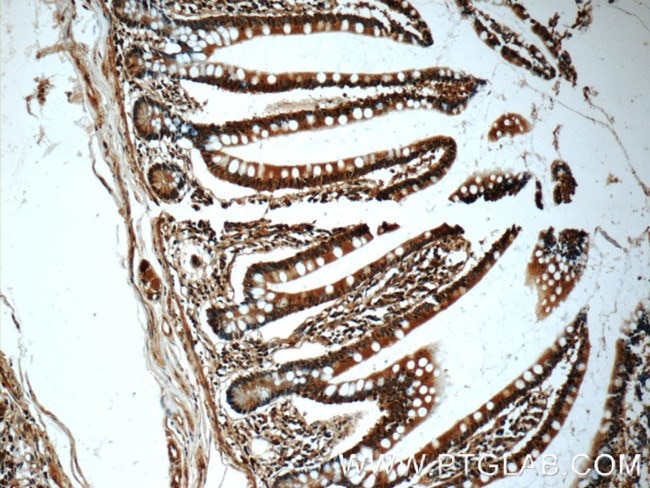
図2. Chk1抗体(カタログ番号:60277-1-Ig、希釈倍率1:50)を使用した、パラフィン包埋ヒト小腸組織スライドの免疫組織化学染色(10倍レンズを使用)
 |
 |
|
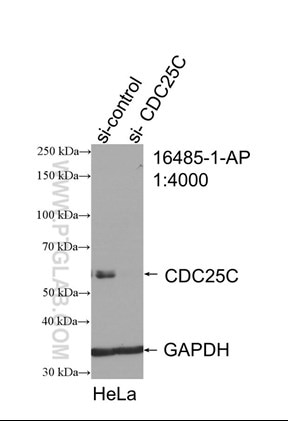
図3. CDC25C抗体(カタログ番号:16485-1-AP、希釈倍率1:4000、室温で1.5時間インキュベート)によるsh-Controlとsh-CDC25CをトランスフェクションしたHeLa細胞のWB(ウェスタンブロット) |
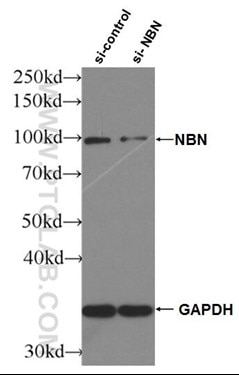
図4. NBS1抗体(カタログ番号:55025-1-AP、希釈倍率1:2000、室温で1.5時間インキュベート)によるsh-Controlとsh-NBS1をトランスフェクションしたHeLa細胞のWB(ウェスタンブロット) |
関連製品
ローディングコントロール抗体
| GAPDH抗体 | |
| カタログ番号:60004-1-Ig | |
|
GAPDHは、多くの種類の細胞で安定して高レベルに発現しているため、ウェスタンブロットでタンパク質のローディングコントロールとしてよく使用されます。この酵素は、解糖、DNA修復、アポトーシス等の細胞で起きる現象に関与します。 |
 |
| Beta Actin抗体(KD/KO validated) | |
| カタログ番号: 66009-1-Ig | |
|
β-アクチンはすべての真核細胞で安定して発現しており、タンパク質の発現レベルがほとんどの実験操作による影響を受けないことから、ローディングコントロールとして一般的に使用されています。 |
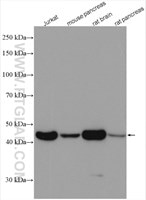 |
プロテインテックのコントロール抗体は、お求めやすい価格¥36,000/150µlで提供しています。
DNA 損傷修復マーカー
|
マーカー名 |
抗体タイプ |
カタログ番号 |
損傷タイプ/ |
|
ATM |
ウサギポリクローナル |
DSB |
|
|
ATR |
ウサギポリクローナル |
SSB |
|
|
ATRIP |
ウサギポリクローナル |
SSB |
|
|
CDC25A |
ウサギポリクローナル |
SSB, DSB |
|
|
CDC25B |
ウサギポリクローナル |
SSB, DSB |
|
|
CDC25C |
ウサギポリクローナル、マウスモノクローナル |
16485-1-AP(図3); 66912-1-Ig |
SSB, DSB |
|
Chk1 |
ウサギポリクローナル、マウスモノクローナル |
25887-1-AP ; 60277-1-Ig(図2) |
SSB |
|
Chk2 |
ウサギポリクローナル |
DSB |
|
|
DDB1 |
ウサギポリクローナル、マウスモノクローナル |
SSB/ NER |
|
|
DDB2 |
ウサギポリクローナル |
SSB/ NER |
|
|
Fen1 |
ウサギポリクローナル |
DSB/ MMEJ |
|
|
H2AX |
ウサギポリクローナル |
DSB |
|
|
Ku70 |
ウサギポリクローナル、マウスモノクローナル |
DSB / NHEJ |
|
|
Ku80 |
ウサギポリクローナル、マウスモノクローナル |
DSB / NHEJ |
|
|
Mre11 |
ウサギポリクローナル |
DSB / HR |
|
|
NBS1 |
ウサギポリクローナル |
55025-1-AP(図4) |
DSB / HR |
|
PCNA |
ウサギポリクローナル、マウスモノクローナル |
SSB / NER |
|
|
POLD1 |
ウサギポリクローナル |
SSB |
|
|
POLD2 |
ウサギポリクローナル |
SSB |
|
|
POLD3 |
ウサギポリクローナル |
SSB |
|
|
POLD4 |
ウサギポリクローナル |
SSB |
|
|
POLE3 |
ウサギポリクローナル |
SSB |
|
|
POLK |
ウサギポリクローナル |
SSB |
|
|
RAD1 |
ウサギポリクローナル |
SSB |
|
|
RAD51 |
ウサギポリクローナル、マウスモノクローナル |
DSB / HR |
|
|
RFC2 |
ウサギポリクローナル |
SSB / NER |
|
|
RFC3 |
ウサギポリクローナル |
SSB / NER |
|
|
RFC4 |
ウサギポリクローナル |
SSB / NER |
|
|
RFC5 |
ウサギポリクローナル |
SSB / NER |
|
|
RPA1 |
ウサギポリクローナル |
SSB、DSB / HR |
|
|
RPA2 |
ウサギポリクローナル |
SSB、DSB / HR |
|
|
RPA3 |
ウサギポリクローナル |
SSB、DSB / HR |
|
|
RPA4 |
ウサギポリクローナル |
SSB、DSB / HR |
表1. DNA損傷マーカーおよびDNA損傷経路に対する抗体のリスト
表中の「損傷タイプ/修復メカニズム」列で使用されている略称:
Double Strand Breaks(DSB)
Homologous Recombination(HR)
Microhomology-Mediated End Joining(MMEJ)
Nucleotide Excision Repair(NER)
Non-Homologous End Joining(NHEJ)
Single Strand Breaks(SSB)
参考文献
1. D. Stein, D. Toiber (2017) “DNA damage and neurodegeneration: the unusual suspect”.
2. T.Lindahl, D. E. Barnes (2000) “Repair of endogenous DNA damage“.